
REVMEDUAS
REVMEDUAS; Vol. 16 No. 1 Enero-Marzo 2026
ISSN 3122-4342
Saúl Armando Beltrán-Ontiveros1, Edgar Iván López-Pulido2, Julio César Villegas-Pineda3, Ana Laura Pereira-Suárez3,4, Daniela Guadalupe Franco-Somera5, José Miguel Moreno-Ortíz6, Kimberly Estefanía Dorantes-Bernal7, Adrián Ramírez-de-Arellano4, Cecilia Rico-Fuentes2*
*Autor de correspondencia: Cecilia Rico Fuentes
Av. Rafael Casillas Aceves No. 1200, C.P. 47620, Tepatitlán de Morelos, Jalisco.
Tel: (331) 271-73-70 E-mail: cilulkal@gmail.com
DOI http://dx.doi.org/10.28960/revmeduas.3122-4342.v16.n1.005
Texto Completo PDFRecibido 18 de septiembre 2025, aceptado 26 de febrero 2025
RESUMEN
Objetivo: El presente proyecto consiste en evaluar la expresión de GLI1 en tejido tumoral y compartimento celular, con diferentes grupos pronóstico-provenientes de tejidos de cáncer de próstata. Materiales y métodos: En este estudio, se utilizó un diseño transversal comparativo, donde se investigó la expresión de GLI1 en tejido tumoral y estromal en tejidos incrustados en parafina con diferentes grupos pronóstico. Tras clasificar las muestras según el puntaje de Gleason (GS), se realizaron análisis de inmunohistoquímica para detectar GLI1 en secciones de tejido, utilizando un sistema automatizado de tinción. Resultados: La expresión de GLI1 demuestra un aumento significativo conforme avanza GS en el tejido tumoral de CaP, lo que sugiere un papel relevante de GLI1 en la progresión tumoral en CaP. Mientras en tejido estromal, la expresión GLI1 se conserva en núcleo y citoplasma conforma avanza GS. Conclusión: la expresión de GLI1 aumenta con el avance de los grados pronósticos en CaP el cual se observó en los citoplasma y núcleo en todos los GS.
Palabras clave: Cáncer de próstata, Vía de Hedgehog, GLI1, Puntaje de Gleason.
ABSTRACT
Objective: This project aims to evaluate the expression of GLI1 in tumor tissue and cellular compartments, with different prognostic groups derived from prostate cancer tissues. Materials and methods: In this study, a comparative cross-sectional design was used to investigate GLI1 expression in tumor and stromal tissue embedded in paraffin, across different prognostic groups. After classifying the samples based on the Gleason score (GS), immunohistochemical analyses were performed to detect GLI1 in tissue sections, using an automated staining system. Results: GLI1 expression shows a significant increase as GS progresses in PCa tumor tissue, suggesting a relevant role of GLI1 in tumor progression in PCa. In stromal tissue, GLI1 expression is maintained in both the nucleus and cytoplasm as GS advances. Conclusion: GLI1 expression increases with advancing prognostic grades in PCa, observed in both the cytoplasm and nucleus across all GS levels.
Keywords: Prostate cancer, Hedgehog pathway, GLI1, Gleason score
Introducción
La próstata es una glándula masculina responsable de producir un líquido que constituye una parte esencial del semen. Cuando las células de esta glándula empiezan a crecer fuera de control, se presenta el cáncer de próstata (CaP). Según las estadísticas, CaP es la causa de muerte más prevalente en la población masculina a nivel mundial.1 El CaP se clasifica según criterios histológicos, clínicos y de diseminación para determinar su gravedad, y asimismo establecer el pronóstico. El sistema o puntuación de Gleason evalúa la agresividad del tumor en un puntaje resultado de una sumatoria que va desde 6 a 10, siendo 6 de bajo grado, 7 intermedio y 8-10 alto. Estableciéndose de esta manera las puntuaciones que van desde 6 (3+3) que representa un tejido sano en composición uniforme hasta 10 (5+5), que representa el grado más avanzado con tumores totalmente indiferenciados y agresivos.2 Es importante mencionar que cada uno de estos grados se asocia con un riesgo diferente de presentar resistencia al tratamiento o recaídas después de una prostatectomía radical que es la extirpación de una parte o la totalidad de la glándula. 3
La vía de señalización Hedgehog (Hh) es una ruta que se ha conservado evolutivamente y juega un papel importante en la regulación de una variedad de procesos fisiológicos y de desarrollo que implican la diferenciación y proliferación de células madre.4 Por ejemplo, durante el desarrollo embrionario, la vía Hh tiene un papel importante en la regulación de los procesos morfológicos complejos, y sus acciones son necesarias para el desarrollo del sistema nervioso central, de las estructuras craneal-facial5, de los tejidos esteroidogénicos entre otras.6
La señalización Hh funciona para activar la transcripción celular de la familia GLI de los factores de trascripción (GLI1, GLI2, and GLI3).7 La señalización canónica clásica se inicia mediante la secreción de péptidos como Shh8, Ihh9 y Dhh10 los cuales reconocen una proteína receptora en la superficie celular (Patched) de las células objetivo.11 La unión de un ligando inicia una inhibición del receptor Smoothened (Smo) el cual altera la fosforilación y los patrones de degradación proteolíticos de GLI2 y GLI3 de un estado represor a un estado activador transcripcional.12 Una vez activados, inducen una expresión de GLI1 lo cual hace que aumente aún más la actividad del estado transcripcional.8 La vía de Hh es también un estímulo proliferativo y la hiperactividad de dicha vía en tejidos adultos puede causar transformaciones neoplásicas y tumores malignos.13
En el caso de pacientes con CaP, la hiperactivación de la vía Hh está asociada con la agresividad, la metástasis y la recaída del paciente. La vía de Hh tiene varios componentes principales: tres homólogos de Hh, PTCH1, el receptor de membrana Smo y los factores de transcripción GLI1, GLI2 y GLI3.14 El factor GLI1 desempeña un papel de activador transcripcional de la vía de Hh uniéndose a la región promotora de genes involucrados en la formación y progresión tumoral.15
En el presente estudio se investigó el papel de la vía Hh en la formación de hiperplasia en células basales de próstata durante la progresión de la tumorigénesis. Para esto se determinó la expresión de GLI1 en tejido tumoral y estromal con diferentes grupos pronóstico de pacientes con cáncer de próstata. Los resultados demostraron que GLI1 aumenta la expresión conforme avanza el grado pronóstico de CaP en tejido tumoral, principalmente en el compartimento nuclear y citoplasmático, mientras que en tejido estromal GLI1 se mantiene en todos los grados pronóstico en el compartimento citoplasmático.
Materiales y métodos
Muestras de tejido
Se trabajó con tejidos incluidos en parafina provenientes de pacientes con diferentes grados pronósticos de CaP, los cuales habían sido examinados y archivados en el Centro de Diagnóstico e Investigación en Patología y Nefropatología. Tras la verificación del diagnóstico y de las categorías pronósticas en Gleason score (GS), las muestras fueron transportadas a nuestro laboratorio y clasificadas en cuatro grupos de grado etiquetados como 2, 3, 4 y 5, de la siguiente manera: GS 7 (3 + 4) (N=9), GS 7 (4 + 3) (N=18), GS 8 (4 + 4) (N=11) y GS 9 (4 + 5) (N=15). La inmunohistoquímica se llevó a cabo en el Laboratorio de Patología Diagnóstica e Inmunohistoquímica de la Universidad de Guadalajara, México. El diseño metodológico aplicado fue transversal comparativo, donde se consideró como criterios de inclusión bloques de parafina con tejido incrustado, excluyendo aquellos bloques que tuviera una poca cantidad de tejido.
Inmunohistoquímica
Los bloques de tejido fijados en formalina e incluidos en parafina se cortaron en secciones de 5 µm, las cuales se tiñeron automáticamente usando el sistema BOND de Leica Biosystems. Se utilizó un anticuerpo primario policlonal contra GLI1 de Santa Cruz (Cat. sc515781) a una dilución de 1:100, seguida de una etapa de incubación con buffer de EDTA durante 15 minutos. Posteriormente se utilizó un buffer diluyente de anticuerpos Diamond de Cell Marque™ Tissue Diagnostics (Cat. 938B-05) y se detectó con Tetrahidrocloruro de diaminobenzidina, DAB mediante Ventana Medical Systems, Roche (Cat. 11718096001).
Realizamos el análisis inmunohistoquímico con un microscopio de luz Axioskop 2 plus (Carl Zeiss, Alemania) acoplado a una cámara digital Coolsnap (Photometrics, Tucson, EE. UU.). Las imágenes fueron documentadas con Aperio de Leica Biosystems, y el análisis densitométrico se realizó usando el software Qupath versión 0.2.3 con un tipo de imagen de campo claro (H-DAB). Los análisis se dirigieron a la tinción nuclear y citoplasmática, y se realizaron utilizando análisis celular y detección de células positivas con un aumento de 40x. Las mediciones se realizaron en tejido tumoral y estromal.
Análisis Estadístico
Las diferencias se consideraron significativas con un valor p ≤ 0.05. Los análisis estadísticos se realizaron utilizando R versión 4.1.2 (R Core Team 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Viena, Austria. URL http://www.R-project.org/, consultado el 13 de enero de 2022). Se utilizaron variables discretas +1, +2, +3, siendo una expresión de menor a mayor respectivamente para la localización nuclear y citoplasmática de las células. Posteriormente, como variables continuas tenemos el porcentaje de células positivas para los tejidos tumorales y estromales, el cual se realizaron a través del programa QuPath versión 3.2.1, finalmente se compararon utilizando un ANOVA de dos colas y una cola con un nivel de significancia de p<0.05.
Declaración de ética
Los estudios que involucraron a humanos fueron aprobados por el Comité de Ética del Centro Universitario de Ciencias de la Salud. Se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. La presente investigación fue aprobada por el Comité de Ética del Centro Universitario de Ciencias de la Salud (Opinión No. CI-01719), donde se consideró la exención del consentimiento informado, basada en las directrices estipuladas en la Guía 10 del CIOMS. Se tomaron en cuenta los siguientes puntos: la investigación no podría haberse realizado sin la exención del consentimiento informado, considerando que nuestro estudio retrospectivo se basó en la recolección de muestras sobrantes de diagnósticos patológicos; no existe riesgo alguno para los participantes, ya que la información personal fue disociada y no es posible identificar a los pacientes. Además, esta investigación tiene un valor social y científico significativo para la población mexicana estudiada.
Resultados
Expresión de GLI1 en el compartimento celular del tejido tumoral y estromal de CaP
Para evaluar la expresión de GLI1, se analizó por inmunohistoquímica el tejido tumoral, se encontró que la expresión se observa en el núcleo y citoplasma en todos los grupos pronósticos (Fig. 1A). Además, mediante la medición por intensidad, se observó una mayor expresión en citoplasma comparado con la expresión nuclear sin presentar diferencias significativas en todos los grados Gleason (Fig. 1B).
Posteriormente, se tiene la expresión de GLI1 en tejido estromal, donde se observó la expresión principalmente en el compartimento citoplasmático en todos los grupos pronósticos (Fig. 2A). Asimismo, en la (Fig. 2B). se presenta la expresión citoplasmática con una intensidad entre +2 y +3 en todos los grupos pronóstico sin presentar diferencias significativas.
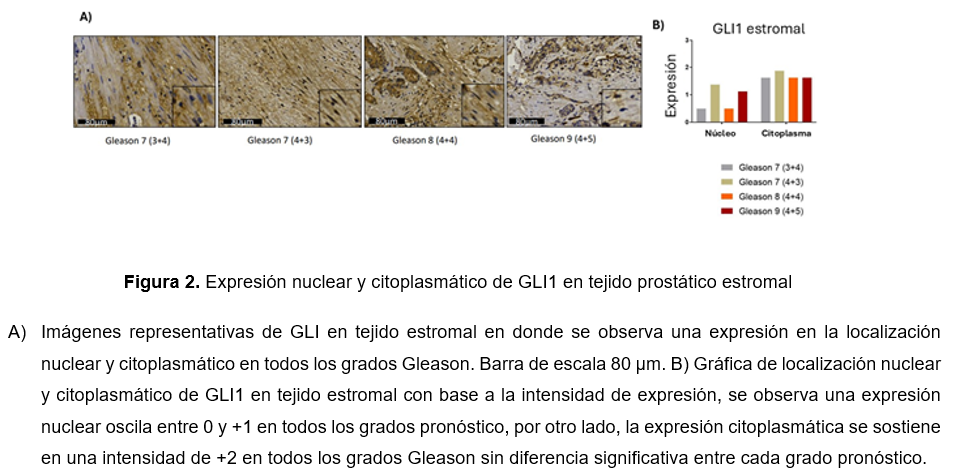
Expresión de GLI1 en tejido tumoral y estromal de CaP
Para obtener una mejor visualización en la expresión de GLI1 en tejido tumoral y estromal, se tomó como referencia el porcentaje de células positivas conforme avanza el grado pronóstico de Cap. En el tejido tumoral podemos encontrar un aumento de expresión entre GS 7 (3 + 4) y GS 9 (4+5) con diferencia significativa de p<0.0001, seguido la expresión entre GS 7 (4 + 3) y GS 9 (4+5) con diferencia significativa de p<0.01 y finalmente la expresión entre GS 7 (3 + 4) y GS 8 (4+4) con diferencia significativa de p<0.05 (Fig.3A). En contraste con tejido estromal, se encontró una expresión sostenida en todos los grados Gleason en el tejido estromal sin diferencia significativa (Fig.3B).
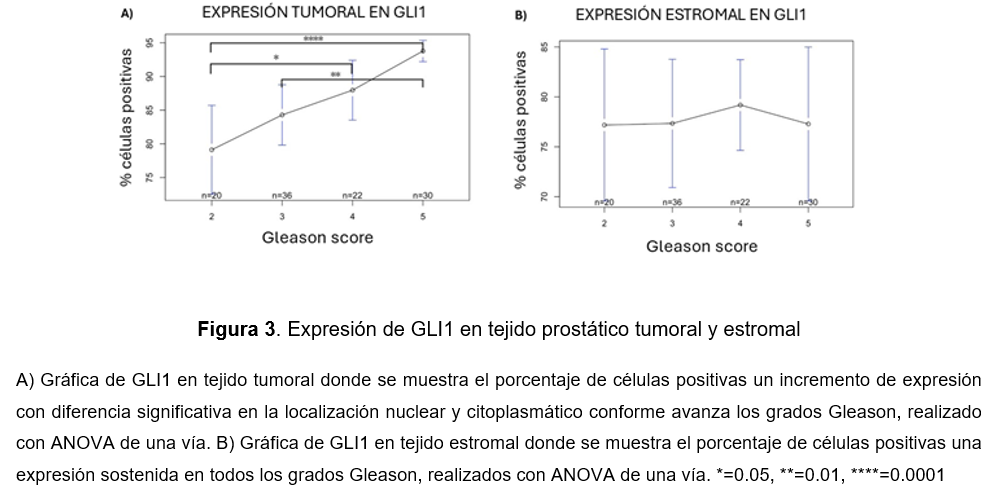
Interacción en la expresión de GLI1 en tejido tumoral y estromal con el compartimento celular en CaP
Para alcanzar un panorama en la participación de GLI1 en todos los grados pronósticos, se tomó como referencia el porcentaje de células positivas conforme avanza el grado pronóstico de CaP, el cual se registró la intensidad como grado 2, 3, 4, 5 como GS 7 (3+4), GS 7 (4+3), GS 8 (4+4) y GS 9 (4+5) respectivamente. Asimismo, se incluyó la expresión nuclear y citoplasmático. Para el tejido tumoral se observó que aumenta la intensidad en la expresión de GLI1 en el compartimento citoplasmático y nuclear (Fig.4A). Posteriormente para el tejido estromal, se observó que no existe una interacción entre la expresión nuclear y citoplasmático conforme avanza los grados de intensidad (Fig. 4B).
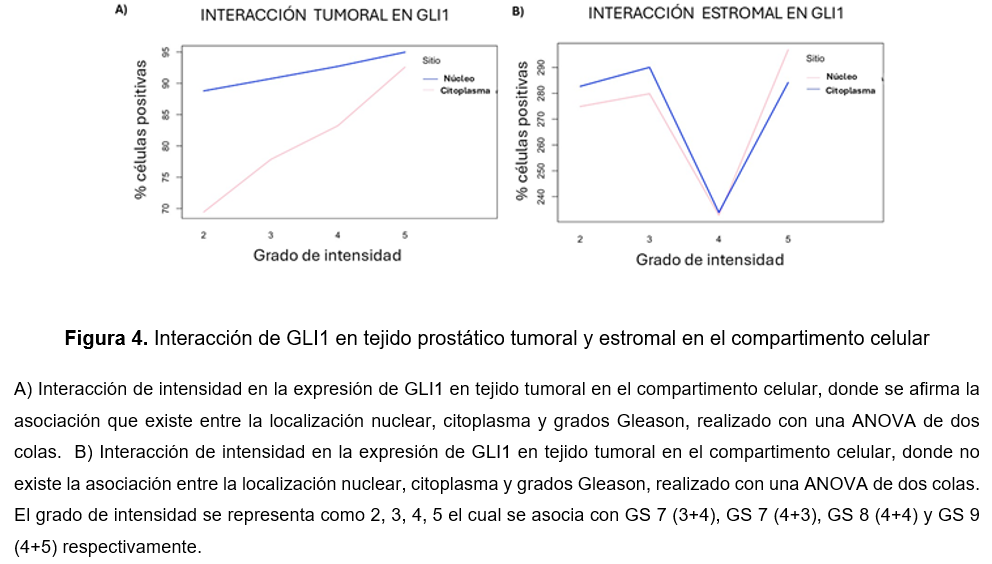
Discusión
Los resultados indican que conforme avanzan el grado de agresividad del CaP, la expresión de GLI1 en el tejido tumoral también aumenta, principalmente en el citoplasma y en menor medida en el núcleo celular, Este aumento en la expresión de GLI1 quizá se asocia con la expresión de HIF-1α, ciclina D1, p27 y p21, que son reguladores positivos del ciclo celular involucrados en un estadio avanzado con metástasis en ganglios linfáticos en CaP.16 Además, en este mismo reporte se encontró que GLI1 regula la EMT a través de la señalización de PI3K/AKT/NF-kB 17, esta expresión se ha visto principalmente en la región citoplasmática en CaP, por lo que, de igual manera, coincide con los resultados obtenidos.
Aunque GLI1 desempeña un papel clave en la activación de cánceres de manera canónica por la vía Hh 18, también es crucial abordar la activación oncogénica no canónica (CMYC, RAS/RAF, TGFβ, etc.). 19 Por ejemplo, en los cánceres gastrointestinales, la sobre activación de GLI1 está impulsada por mutaciones en KRAS/BRAF. 20 Recientemente se ha sugerido que el GLI1 oncogénico progresa durante la carcinogénesis del colon 21,22 y en la enfermedad metastásica mientras que, en el tejido colónico normal, la vía Hh-GLI está estrictamente involucrada en la diferenciación. 23
Otro ejemplo, se ha encontrado en cáncer de pulmón de células no pequeñas, en donde GLI1 contribuye a la angiogénesis mediante la vía de señalización Shh/GLI1 en tejido tumoral.24 Dicho estudio identificó a GLI1 como un regulador crítico de la interacción entre las células de cáncer de pulmón de células no pequeñas y las células vasculares.25 GLI1 en las células de cáncer de pulmón de células no pequeñas promueve la motilidad de células endoteliales necesaria para la angiogénesis mediante la promoción de la expresión del factor transformador de crecimiento β (TGFβ). 26
Respecto al tejido estromal, la expresión de GLI1 en el citoplasma y en el núcleo celular se mantiene sin tendencia en todos los grados pronósticos, lo que sugiere la influencia del microambiente tumoral en la progresión del CaP. Se ha demostrado que en vías de señalización Shh-GLI1 es esencial para determinar los niveles de expresión de los marcadores de células madre de CaP, ya que aumenta el número y tamaño de los esferoides prostáticos in vitro.27 Por el contrario, la inhibición de la señalización de GLI1 en el estroma suprime la diferenciación de las células basales hacia células luminales, asimismo limita la capacidad secretora de los túbulos prostáticos por lo que bloquea la regeneración del tejido prostático in vivo. 28
A nivel molecular, la supresión de la señalización GLI1 aumenta el factor de señalización en TGFβ en las células estromales de CaP. La elevación de los niveles de TGFβ inhibe la formación de esferoides prostáticos, lo que sugiere que un eje de señalización GLI1-TGFβ en el estroma regula la actividad de las células progenitoras epiteliales, por lo que es tiene potencial de renovación en ambos compartimentos, epitelial y estroma en CaP. 29
En otros cánceres, en un estudio previo de Rebekka y col. destacan el papel crucial de la activación de GLI1 las células estromales en la patogénesis de la fibrosis de médula ósea.30 Se encontró que puede ser activada por plaquetas o megacariocitos atípicos a través de Cxcl4, lo que desencadena procesos de diferenciación miofibroblástica, reprogramación metabólica y disminución de CXCL12.31 Además, se propone que las proteínas GLI1 en estas células pueden activarse de manera independiente de la señalización canónica de Hh, posiblemente mediante la vía PI3K/AKT, lo que explicaría la variabilidad en las respuestas de los pacientes con fibrosis de médula ósea. 30
En conclusión, estos hallazgos subrayan el papel de GLI1 en el tumor y en estroma consolidándolo como un regulador clave en la progresión tumoral y un posible blanco terapéutico en el manejo de CaP. Asimismo, las interacciones que existe en el compartimento celular como las interacciones en el tejido tumoral y estromal, lo cual nos abre un amplio panorama interesante por explorar.
Conflicto de interés
Los autores no declaran conflicto de interés
Agradecimientos
CONAHCYT, Ciencia Básica y de Frontera 2023-2024 (CBF2023-2024-2702)
Referencias